Autor: Alvaro Segura Monge
Profesor de la Universidad de Costa Rica
La ¨Sigatoka negra¨ (SN) también conocida como BLSD, es causada por el hongo ascomiceto Pseudocercospora fijiensis M. Morelet-Deighton, anteriormente denominado como Mycosphaerella fijiensis M. Morelet (Arango-Isaza et al., 2016). Este hongo ha sido durante mucho tiempo una limitación importante para la producción de banano y plátano induciendo una defoliación severa y afectando el rendimiento en la mayoría de las zonas tropicales y subtropicales donde se cultivan bananas y plátanos (Carlier et al., 2000). Según Guzmán et al., (2019) en cultivares de banano susceptibles a la ¨Sigatoka negra¨, el impacto por la reducción del follaje, puede causar una reducción entre el 20% y 40% en el peso de la fruta y un daño del 100% en la calidad de exportación.
El propósito de este trabajo es hacer una revisión de los procesos fisiológicos en la relación patógeno-planta de banano, que conducen a la disminución del peso del racimo de banano por efecto de la incidencia y severidad de P.fijiensis.
Antecedentes del efecto del área foliar sobre el peso del racimo de banano:
El peso del racimo del banano está condicionado por el potencial fotosintético de la planta, es decir la capacidad de la planta de reducir CO2 a través de un proceso, donde participan la radiación solar, el suministro de agua y nutrimentos, entre otros componentes. La capacidad de la planta para utilizar las sustancias que proveen energía, denominados fotoasimilados, va a depender del balance entre el carbono reducido, vía fotosíntesis, y el carbón oxidado, a través de la respiración y/o la fotorespiración (en plantas C3 como el banano). La diferencia entre ambos procesos es denominada como Fotosíntesis neta (Fn), y constituye la fuente de carbono que la planta usa para sustentar la producción de biomasa. Durante el crecimiento y desenvolvimiento de la planta de banano se produce un incremento del volumen de materia seca, la cual es compartimentada en diferentes secciones, llámese, hojas, raíces, rizoma, pseudotallo, hijos de sucesión y racimo (Segura et al., 1995 b).
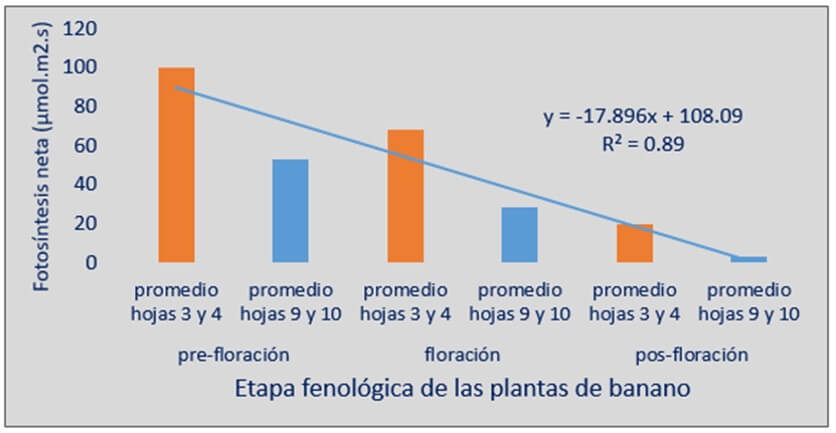
Segura et al., (2005 a) determinaron el efecto de la edad de las hojas de plantas de banano de tres cultivares ¨Grande Naine¨, ¨Williams¨ y ¨Valery¨, sobre la fotosíntesis neta foliar (Fn) antes de la floración, a la floración y a pos-floración en plantas sanas por efecto de la ¨Sigatoka negra¨. Se encontró que la fotosíntesis neta disminuyó, desde las hojas apicales a basales. Las tres primeras hojas formadas, presentaron las mayores tasas fotosintéticas. Este comportamiento lo mostraron todos los cultivares, con pocas variaciones en los valores puntuales de Fn por hoja. La caída en la Fn fue igual en todos los estadios fenológicos evaluados; sin embargo, los valores de Fn fueron inferiores en todas las hojas conforme la planta pasó de estadios juveniles a pre-floración, adulta pre-floral a pos-floración (Figura 1)
|
Figura 1. Relación entre edad de las hojas y la tasa de fotosíntesis neta promedio para los cultivares “Grande Naine”, “Williams” y “Valery” (Grupo Cavendish, AAA). Fuente: Segura et al., 2005 |
La disminución de la fotosíntesis neta foliar que ocurrió con la edad de las hojas fue más evidente en la medida que la planta alcanzó estados fenológicos más avanzados. Una razón muy importante para justificar este comportamiento proviene del hecho que en la medida que avanzan los cambios fenológicos en la planta, la floración y el llenado del fruto se convierten en ¨drenos¨ o ¨sumideros¨ de energía prioritarios del carbono reducido en las hojas a través del proceso de fotosíntesis, por lo tanto, la tasa de fotosíntesis neta tiende a disminuir. Por otro lado, se sugiere, que, desde el punto de vista energético, la planta puede utilizar otros reservorios de energía y con ello compensar la caída o dilución de la tasa fotosíntesis neta foliar que provoca el efecto dreno tanto del racimo, como del hijo de sucesión en crecimiento (Segura et al., 2005 b). Los resultados indican que las hojas más basales de la planta realizan una menor contribución de carbono a la planta.
Dado que la tasa fotosintética disminuye con la edad de las hojas, es posible sugerir que más allá de la hoja 9 a la floración, estas no solo no aportan fotoasimilados en forma significativa al resto de la planta, particularmente al racimo, sino que también estas hojas basales podrían convertirse en una barrera que obstaculice la recepción de la luz directamente al racimo, y con ello limitar una eventual fijación de carbono directamente del fruto, toda vez que, la cáscara, de los frutos contienen, clorofila y estomas (Moser et al., 2009, Moser et al., 2012). De momento no existe información disponible que indique que los frutos de banano puedan fijar carbono; pero hay evidencia que ocurre en otros frutos (Blake, 1990).
Adicionalmente el acúmulo de ceras epicuticulares que se observa al aumentar la edad de las hojas del banano, puede dificultar el intercambio de CO2 a través de las estomas y limitar su potencial de reducción de carbono (Freeman y Turner, 1985).
Segura y Valle (2005) estudiaron el efecto de la deshoja artificial selectiva al momento de la floración de plantas de los cultivares ¨Grande Naine¨, ¨Williams¨ y ¨Valery¨, con un muy buen manejo de la ¨Sigatoka negra¨, sobre algunos componentes del rendimiento y calidad de los frutos al momento de la cosecha. Se mantuvieron plantas con 7, 9, 11, 13 y 15 hojas retenidas. No se encontraron diferencias estadísticas entre los niveles de defoliación respecto a la altura y circunferencia del hijo de sucesión a la cosecha, en el número de manos, en el peso del racimo, ni tampoco en el grosor y longitud de los dedos de las manos sub-basal y apical; sin embargo, se observó que las plantas con menor número de hojas retenidas, mostraron una menor duración del período comprendido entre la floración y la cosecha, lo cual provocaría un aumento en la velocidad de retorno. Resultados similares fueron hallados por Vargas et al., (2006). Por su parte Vargas et al. (2005, 2006), Saborío y Granados (2005) y Rodríguez et al., (2006), obtuvieron resultados similares en los estudios sobre deshoja controlada.
Segura et al. (2005 b) al evaluar el crecimiento y particionamiento de la biomasa de los cultivares ¨Grande Naine¨, ¨Williams¨ y ¨Valery¨, partiendo de plantas de cultivo de tejidos; observaron que durante la etapa juvenil hay un incremento considerable de la biomasa de hojas y un aumento paulatino en la del pseudotallo y el rizoma, con pocas variaciones en la de las raíces. A partir de la floración se observó una disminución de la biomasa foliar y del rizoma, un incremento en la del hijo de sucesión y del racimo, y aproximadamente dos semanas después de la floración una notable caída en la biomasa acumulada en el pseudotallo. Así desde la floración a la cosecha la biomasa de las hojas y el pseudotallo pasaron de un 40 y 30% hasta un 10 y 20%, respectivamente. Se presume que las exigencias energéticas del racimo y del hijo de crecimiento son particularmente suplidas por la biomasa acumuladas en las hojas y el pseudotallo; sin embargo, al ser el banano una planta perenne de sucesión, la biomasa aportada por el pseudotallo tiene mucho significado en la sostenibilidad de la producción en las generaciones sucesivas, ya que a partir de la emisión floral las hojas, no hay nueva producción de hojas y como se mencionó, estas tienden a reducir su tasa de reducción de carbono.
Los pseudotallos, y en menor grado los rizomas de las plantas de banano, son reservorios energéticos y de nutrimentos de mucha importancia para la planta, de manera que cuanto mayor es la circunferencia de la planta madre de plantas de los cultivares ¨Grande Naine¨, ¨Williams¨y ¨Valery¨ (grupo Cavendish,
AAA), medidas a la floración, mayor es el peso del racimo (Segura y González, 2005 datos sin publicar). En adición al importante aporte de fotoasimilados y nutrimentos que proporcionan las hojas para el crecimiento del racimo, hay una coparticipación de otros reservorios energéticos, que hacen a la planta menos prescindible energéticamente de las hojas para atender sus necesidades, particularmente para determinar el peso y la calidad del racimo.
De acuerdo con Segura et al., (2005) la biomasa acumulada en el pseudotallo de plantas de banano (grupo Cavendish, AAA) después de la cosecha de la primera generación (R0), constituye el 20% de la materia total de la planta. Por su parte Medina (2007), determinó que este reservorio representó el 27% del total de la biomasa de una planta cosechada de quinta generación (R5).
Impacto de la ¨Sigatoka negra¨ sobre el peso del racimo:
El efecto de la enfermedad en la producción y calidad, es más evidente en suelos de baja fertilidad o en condiciones de deficiencia nutricional (Cuadro 1).
|
Cuadro. 1 Efecto de la “Sigatoka negra” sobre algunas variables productivas del banano “Grande naine” (Subgrupo Cavendish AAA) bajo condiciones de baja y alta fertilidad del suelo. Costa Rica (Guzmán, M. y Villalta, R. 2015. Datos no publicados) |
||||
|
Suelo de baja Fertilidad |
||||
|
|
Peso racimo (Kg) |
Numero de manos |
Diámetro del dedo central de la última mano (mm) |
Longitud del dedo central de la última mano (cm) |
|
Con control químico |
24.1 |
6.7 |
32.8 |
20.5 |
|
Sin control químico |
16.9 |
6.4 |
29.3 |
18.2 |
|
Diferencia % |
-29.9 |
-9.6 |
-10.7 |
-11.2 |
|
(P<0.0001) |
** |
ns |
** |
** |
|
Suelo de alta fertilidad |
||||
|
|
Peso racimo (Kg) |
Numero de manos |
Diámetro del dedo central de la última mano (mm) |
Longitud del dedo central de la última mano (cm) |
|
Con control químico |
29.2 |
8.0 |
33.3 |
19.8 |
|
Sin control químico |
21.9 |
7.8 |
30.0 |
18.1 |
|
Diferencia % |
-25 |
-2.5 |
-9.9 |
-8.6 |
|
Con control químico |
** |
ns |
** |
** |
El principal efecto de P. fijiensis sobre el peso del racimo del banano es quizá atribuible a una reducción de la tasa fotosintética foliar por acción directa del daño provocado por las fitotoxinas y la producción de especies reactivas de oxígeno (ROS), que dañan severamente a los tejidos foliares (Segura, 2020).
Según Rodríguez y Cayón (2008), al incrementarse la severidad de la ¨Sigatoka negra¨, las hojas enfermas mostraron alteraciones en todas las variables fisiológicas evaluadas, determinándose una pérdida notable de biomasa y de pigmentos fotosintéticos desde los estadios iniciales de infección, afectando directamente procesos fisiológicos básicos como fotosíntesis y transpiración. La resistencia y la actividad estomática, se alteraron en relación directa con el aumento de la severidad de la enfermedad. Se observó un daño severo de las cavidades estomáticas, causado por la penetración del hongo y por la extrusión del inóculo para nuevas infecciones, todo lo cual provocó una disminución de la tasa de fotosíntesis neta, lo cual es coincidente con los resultados informados por Hidalgo et al., (2006).
Como consecuencia del daño directo del patógeno a la estructura fotosintética foliar, las concentraciones de almidón y azúcares disminuyeron; sin embargo, presumiblemente también indirectamente, por el consumo de estas fuentes de energía por parte del hongo para sustentar su actividad. La disminución de la síntesis de almidón y azúcares a nivel foliar provocaría una eventual reducción del peso de la fruta, toda vez que esta última requiere del aporte de fotoasimilados que le provee las hojas para su normal crecimiento.
Las unidades de calor, grados día (dd) o madurez fisiológica, se refiere al efecto del acúmulo de calor o suma de la temperatura promedio que ocurre desde la floración, hasta que la fruta se cosecha, por encima de un valor de referencia que son 14°C. Este criterio tiene mucha validez toda vez que existe una variación en la temperatura a través del año, que independientemente de la edad cronológica, impacta el crecimiento y madurez fisiológica de la fruta. Las variaciones de temperatura son más amplias en condiciones subtropicales; sin embargo, aún en condiciones tropicales existen diferencias. Se pueden cosechar frutas con igual edad cronológica través del año, pero con diferente edad fisiológica. Dado que el banano es una fruta climatérica (produce auto-catalíticamente etileno después de alcanzar la madurez fisiológica, induciendo la maduración de la fruta o pérdida de vida verde) los criterios para su cosecha deben ser manejados con mucha precisión para optimizar la relación entre el peso y la duración de la vida verde, toda vez que para efecto de exportación el banano tiene que llegar al mercado verde.
Según Jullien (2008) cuando el llenado de la pulpa no se ve obstaculizado por ningún estrés en la planta, los bananos cosechados a 900 grados día en el umbral de 14 °C, alcanzan un diámetro comercial con una vida verde de aproximadamente 30 días a 20° C. (Ganry y Meyer, 1975).
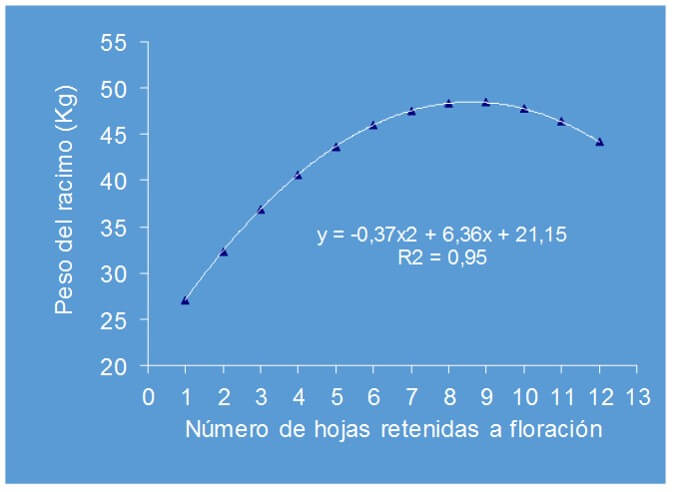
Robinson et al., (1992) al evaluar el efecto de la remoción artificial de las hojas de plantas de banano (grupo Cavendish, AAA) al momento de la floración, sobre el peso del racimo en condiciones de África del Sur, encontraron que el peso de la fruta tendió a incrementarse en la medida que aumentó el número de hojas retenidas, de manera tal que al disponer entre 8 y 9 hojas, se alcanzó el mayor peso; no obstante al aumentar el número de hojas retenidas por encima de estos valores, el peso del racimo tendió a ser menor (Figura 2). Por su parte Daniells et al. (1994) mencionan que las plantas no comprometen el peso del racimo cuando retienen a la floración no menos de 10 hojas.
|
Figura 2: Efecto del número de hojas retenidas a la floración sobre el peso del racimo de banano (grupo Cavendish, AAA). Fuente: Robinson et al., (1992) |
Ramsey et al., (1990) determinaron que en relación a Pseudocercospora musicola (Sigatoka amarilla), las plantas cosechadas con menos de cinco hojas efectivas tuvieron el menor peso de fruta. Igualmente encontraron una correlación negativa altamente significativa entre la severidad de la enfermedad y el número de hojas efectivas a la cosecha. Vargas et al., (2009), al estimar el efecto de la defoliación controlada a la floración de plantas de banano (grupo Cavendish), sobre algunas variables del daño producido por la ¨Sigatoka negra¨, no obtuvieron diferencias en el peso del racimo y el diámetro del dedo al permanecer retenidas no menos de 7-8 hojas a la floración, siempre que a la cosecha hubieran no menos de cinco hojas efectivas. Además, observaron que, al permanecer no menos de 7-8 hojas retenidas, la severidad de la enfermedad no se redujo en la planta sujeta a defoliación controlada, pero si hubo una tendencia a disminuir en las plantas de las siguientes generaciones, impactando positivamente en la reducción del potencial de inóculo de la plantación y en el incremento de la velocidad de retorno (Vargas et al., 2009).
Castelan et al., (2012) estudiaron el efecto de la severidad de la ¨Sigatoka negra¨ sobre las dimensiones y el peso del racimo del cultivar ¨ Nanicão¨ (Cavendish, AAA) cosechado a 700 y 960 grados día, respecto a plantas con un manejo óptimo de la enfermedad. Al comparar la fruta ¨joven¨ cosechada a los 700 grados día, las plantas enfermas tuvieron un período de floración a cosecha de 137 días, respecto a 147 días en las plantas testigo. Es decir hubo un acortamiento del período en las plantas severamente afectadas por la enfermedad y una diferencia de 5% en el peso a favor del testigo. A los 960 grados día, el perído de floración a cosecha fue de 166 días en las plantas testigo y el 100 % de las plantas afectadas por la enfermedad maduraron en el campo. La enfermedad provocó un acortamiento del tiempo entre floración y cosecha, una disminución del peso del racimo, lo cual es coincidende con lo encontrado por Mobambo et al., (1993).
Posteriormente, Castelan et al. (2013) evaluaron el efecto de la severidad de la ¨Sigatoka amarilla¨ sobre las dimensiones de la fruta de banano (¨Grande Naine¨) al cosechar la fruta a 900 grados día, y obtuvieron una relación lineal negativa entre el valor acumulado de la severidad de la enfermedad entre la floración y la cosecha (área bajo la curva), respecto al peso y ligeramente en relación al diámetro de la fruta; pero ninguna diferencia en relación a la longitud del dedo.
Chillet et al. (2009) no encontraron diferencias significativas en el diámetro de los dedos de banano (Cavendish), al comparar plantas con una muy baja y alta infección de P.musicola, y una edad fisiológica de cosecha de 900 grados día. Según lo autores aparentementre el grado de la defoliación inducida por la enfermedad, no fue lo suficientemente crítica para a afectar el crecimiento de la fruta. Sin embargo, este efecto fue inesperado y difiere del efecto de otro tipo de estreses sobre el crecimiento del fruto del banano, tales como la deficiencia de minerales, la escasez de agua (exceso o escasez) y la fuerte defoliación, que muestran un impacto muy claro en la tasa de crecimiento del diámetro de la fruta (Chillet et al., 2006).
Estudios posteriores realizados por Chillet et al., (2013), sobre el efecto de dos niveles de deshoja (no eliminando las hojas enfermas un mes antes de la cosecha y eliminando las cinco hojas más viejas afectadas por la enfermedad) en plantas de ¨Grande Naine¨ afectadas por P.musicola; observaron que los dedos de los racimos cosechados a 900 grados día, fueron significativamente más pesados en las plantas con menor severidad de la enfermedad respecto a las más afectadas. Al comprar los niveles de deshoja de plantas con baja y alta severidad de la enfermedad, el peso de los dedos fue mayor en las plantas que no se les retiró las hojas antes de la cosecha. No encontró diferencias en la circunferencia de la fruta entre niveles de severidad de la enfermedad; sin embargo, si las hubo en la circunferencia, y fue mayor significativamente en las plantas que nos les fueron removidas las hojas.
Por consiguiente, los frutos de las plantas con un alto nivel de infección presentan el mismo volumen (diámetro y longitud) que los de las plantas con un bajo nivel de infección, pero con un peso inferior, lo que probablemente indica un fruto de menor densidad. Por lo tanto, es posible que el llenado de la fruta se viera afectado por la severidad de la enfermedad.
Contario a lo informado por M. Guzmán y R. Villalta, 2015 (resultados no publicados) para el caso de la ¨Sigatoka negra¨, la enfermedad si afectó el diámetro y el peso del racimo; no obstante, los autores no mencionan la edad fisiológica a la cosecha, como si lo indican Chillet et al. (2009), Castelan et al. (2012) y Chillet et al., (2013). Este es un tema sensible, ya que el diámetro del dedo central de la segunda mano basal se utiliza como criterio convencional para confirmar el momento apropiado de cosecha; el cual puede variar estacionalmente según las variaciones de temperatura, pero la edad fisiológica al depender de las unidades calóricas, siempre se alcanza a un valor específico, que según se indicó, en los trópicos es de 900 grados día.
Los resultados obtenidos en este estudio revelaron que los bananos cosechados a la misma edad fisiológica (700 dd o 960 grados día) en parcelas con una alta tasa de infestación por ¨Sigatoka amarilla¨ o ¨Sigatoka negra¨, tenían una vida verde más corta y un pico climatérico más fuerte (producción de CO2 y C2H4) que los bananos cosechados de parcelas con baja severidad de la enfermedad. (Castelan 2013)
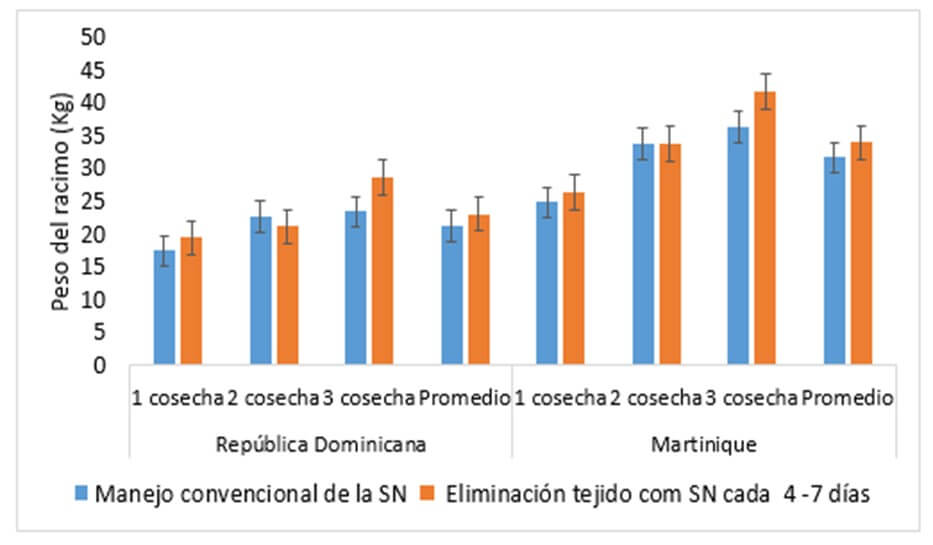
Guillemet et al., (2018) evaluaron dos sistemas de manejo de la ¨Sigatoka negra¨ en República Dominicana, (700 mm de lluvia anual y una temperatura promedio de 25,8 °C) y en Martinica (2100 mm de precipitación anual y una temperatura anual promedio de 25,7 °C) en plantaciones del cultivar ¨Grande Naine¨ por tres ciclos de cultivo. El control recibió un manejo químico convencional cada 3 o cinco semanas, complementado con la eliminación del tejido foliar necrótico con una frecuencia de 7 a 15 días. El segundo tratamiento, no incluyó control químico, pero si la eliminación del tejido necrótico causado por la enfermedad cada 4 a 7 días dependiendo de la evolución de la infección.
Al evaluar el área foliar de las plantas, se encontró que en República Dominicana el primer año, ambos tratamientos mostraron áreas foliares similares a la cosecha, pero el segundo y tercer año, las plantas con manejo convencional mostraron una mayor área foliar que el tratamiento que recibió solo ¨cirugía´ de tejidos. En Martinica, por el contrario, en los tres ciclos, las plantas sin tratamiento químico, llegaron a cosecha cercanas a cero hojas. En ambos lugares, las plantas mostraron un área foliar similar al momento de la floración. Los racimos fueron cosechados a una edad fisiológica entre 800 y 1000 grados día, no observándose diferencias en el peso del racimo en ninguna de las cosechas y tampoco en la duración de la vida verde (Figura 3). Según los autores en ambos tratamientos, las hojas presentes en el momento de la floración, quizá fueron capaces de seguir suministrando fotoasimilados, de manera que no comprometieron el peso del racimo y sugieren, además, la presencia de una fuente compensatoria de energía, en adición a la proveída por la carencia de hojas.
Se sugiere que esta fuente accesoria de energía, pudo haber sido el pseudotallo y en menor grado el rizoma, según lo señalado por Segura et al., (2005 a) y Segura et. al, (2005 b).
Según Guillemet et al., (2018), los resultados obtenidos con ¨Sigatoka negra¨, coinciden con los obtenidos con la ´Sigatoka amarilla¨ por Chillet et al., (2013), mostrando que cuando se practica la eliminación regular de los estadios necróticas del patógeno de las hojas, el impacto de la enfermedad sobre el peso de la fruta es bajo, siempre que se coseche a una madurez fisiológica óptima.
Guillemet et al., (2018) sugieren la importancia de promover la formación de una buena área foliar antes de la floración de las plantas, para permitir su adecuado crecimiento. Por otro lado, durante la fase vegetativa, se formarán otras reservas de energía que contribuyen a suplir las necesidades del racimo. El número de hojas en el momento de la cosecha parece un criterio empírico para indicar el potencial de cosecha de la fruta exportable. Así, al poseer apropiadas prácticas agronómicas, realizar la cosecha en el momento fisiológico óptimo, la eliminación regular de hojas u secciones de hojas enfermas; la ausencia de hojas en el momento de la cosecha, no parece crítica para la producción de frutos compatibles con las normas de exportación.
El estudio en referencia no muestra información sobre la evolución de la severidad de la enfermedad, y considerando que las condiciones de lluvia de las regiones donde se realizó el estudio, son quizá ¨benevolentes¨, en relación a las condiciones que prevalecen en otros lugares, dígase en la región caribe de Costa Rica, hay que tener mucho cuidado con la interpretación y aplicación de esta información. Es importante rescatar de estos estudios, la aparente relación entre los factores determinantes del bajo peso y el acortamiento de la vida verde de la fruta, y la presencia del patógeno en las lesiones necróticas de las hojas.
|
Figura 3: Efecto de dos tipos de manejo de la ¨Sigatoka negra¨ sobre el peso del racimo. Manejo convencional: Se aplicó fungicidas cada 3 semanas en República Dominicana, y cada 3 a 5 semanas en Martinica, con fungicidas sistémicos y protectores aplicados en emulsión. Las partes de la hoja dañadas por ¨Sigatoka negra¨ fueron retiradas cada 7 a 15 días. Eliminación del tejido con SN: Sin fungicidas, solo ¨cirugía¨ de tejidos necróticos tan pronto aparecían cada 4 a 7 días. La sobreposición de líneas indica que son iguales al 0.05% de probabilidad. Fuente: Guillermet et al. (2018) |
Según Guzmán et al., (2019) en Costa Rica se recomienda que las tres hojas más antiguas se eliminen en la primera semana después de la aparición de la bellota floral, dejando la planta con nueve a diez hojas funcionales (Figura 4). El uso continuo de la práctica durante más de 3 años en una plantación de 300 hectáreas que cultiva un cultivar Cavendish mejoró el control de enfermedades y redujo la cantidad de aceite mineral necesario en las medidas de control químico complementarias (Guzmán, 2019). La defoliación controlada se ha implementado con éxito en más de 3000 hectáreas de plantaciones bananeras en Costa Rica (G. Murillo y M. Guzmán, Costa Rica, 2016, comunicación personal).

|
Figura 4. Efecto de la deshoja controlada en banano a la floración para reducir el inóculo de la Pseudocercospora fijiensis. Antes y después de la deshoja (fotografías superiores). Superficie inferior de las hojas eliminadas y senescentes después de su eliminación de la planta, mostrando lesiones detenidas del hongo y en menos número (fotos: M. Guzmán, CORBANA). Fuente: Guzmán et al., (2019). |
De acuerdo con Guzmán et al., (2019) si el tejido necrótico foliar provocado por la ¨Sigatoka negra¨ permanece en la planta, la producción de pseudotecios continúa y las ascosporas pueden ser liberarse hasta por 22 semanas; pero en las lesiones de las hojas eliminadas que caen al suelo, la liberación de ascosporas es de 6 semanas o menos. Además, las esporas liberadas por el tejido foliar en el suelo tienen una probabilidad reducida de alcanzar las hojas jóvenes susceptibles de las plantas.
Marín et al., (2003) mencionan que la reducción de los niveles de inóculo por la eliminación temprana de las hojas de banano con necrosis extensa causada por manchas coalescentes y la escisión de pequeñas áreas de tejido foliar necrótico, prácticas conocidas como 'poda de hojas', "defoliación', ´despunte', o 'cirugía', (según sea el tejido eliminado), contribuyen significativamente a la reducción del inóculo de P.fijiensis y a una mejorar la sanidad de las plantaciones.
Fisiológicamente se justifica la pérdida del peso de la fruta por efecto de la severidad de la ´Sigatoka negra´ por la reducción del área foliar de las plantas, de la tasa fotosintética y de los fotoasimilados (carbohidratos totales, minerales etc.) que son requeridos por el racimo para su crecimiento.
Comentarios finales:
- Las plantas de banano tienen la habilidad de utilizar varios reservorios de energía para abastecer sus necesidades, particularmente en lo que se refiere al racimo. Las hojas tienen una funcionalidad parcial en su capacidad para reducir carbono, de manera que, al envejecerse, disminuyen su fotosíntesis neta. Por otro lado, aún las hojas más jóvenes, disminuyen su capacidad de reducir carbono después de la floración por el efecto ¨dreno’ o ´sumidero¨ de fotoasimilados que provoca el racimo en crecimiento. Esto es, es mayor la demanda de fotoasimililados, que la capacidad de las hojas para proveerlo. Por estas circunstancias, particularmente el pseudotallo se convierte en una fuente accesoria de energía muy importante para la planta, por ser esta una especie perenne.
- Por efecto de la producción de fitotoxinas por parte de Pseudocercospora fijiensis se provocan daños irreversibles en la estructura y funcionalidad de los cloroplastos, provocando su deterioro y capacidad de reducir y acumular fotoasimililados. Como consecuencia directa, se reduce notablemente la fuente de energía que la planta necesita para abastecer sus necesidades vitales, donde el peso del racimo es afectado de forma significativa al aumentar la severidad de la ´Sigatoka negra¨.
- La ¨Sigatoka negra¨ causa una disminución del período de floración a cosecha, es decir un acortamiento del ciclo por el efecto estrés, lo cual es común en plantas sometidas a diversos tipos de tensiones sean estas de naturaleza biótica u abiótica.
- La menor edad fisiológica de las futas de plantas afectadas por la ¨Sigatoka negra¨ ocasiona una maduración temprana y un acortamiento del período pre-climatérico o de vida verde.
- Se informa que por efecto de la enfermedad hay una disminución marcada del peso del racimo, sin que esto implique una reducción en la circunferencia de los dedos, lo cual puede ser causado por una pérdida de la densidad en la pulpa.
- En apariencia existen factores de naturaleza fisiológica asociados a las lesiones del hongo, que se transmiten a la fruta, de manera tal que al implementar medidas de eliminación del tejido foliar afectado, en adición a disminuir el potencial de inóculo del hongo, pueden contribuir a reducir los efectos de la enfermedad en términos del peso del racimo. No hay sin embargo al momento, evidencia experimental que compruebe estas conjeturas.
- La eliminación de las cuatro hojas más viejas al momento de la floración constituye una buena oportunidad para reducir el potencial de inóculo de la enfermedad, sin detrimento de la productividad, toda vez que son hojas que contribuyen con muy poco desde el punto de vista fotosintético.
Agradecimiento: Al Dr. Douglas Marín PhD., por la revisión del manuscrito y por sus acertadas recomendaciones.
Cita correcta de este artículo
Segura, M. A. 2021. Procesos Fisiológicos Involucrados en el Daño Producido por Pseudocercospora fijiensis en las Plantas de Banano (Musa spp): Efecto sobre el Follaje y Disminución del Peso del Racimo (parte 3). Serie Frutales, Núm. 74. Artículos técnicos de INTAGRI. México. 15 p.
Literatura consultada
- Arango, I R.E.; Diaz, T. C.; Dhillon, B.; Aerts, A.; Carlier, J.; Crane, C.F. 2016. Combating a Global Threat to a Clonal Crop: Banana Black Sigatoka Pathogen Pseudocercospora fijiensis (Synonym Mycosphaerella fijiensis) Genomes Reveal Clues for Disease Control. PLOS Genet. 12(8).
- Blanke, M.M. 1990. Photosynthesis in Subtropical Fruits-Review Paper. Acta Horticulturae. 275: 435-439.
- Carlier, J.; Foure´, E.; Gauhl, F.; Jones, D.R.; Lepoivre, P.; Mourichon, X. 2019. Fungal Diseases of the Foliage: Sigatoka leaf spots. In: Jones, D.R., editor. Diseases of Banana, Abaca´ and Enset. Wallingford, Oxon OX10 8DE UK: CABI Publishing.
- Castelán, F.P.; Saraiva, L.A; Lange, F.; Bellaire, L.D.L.; Cordenunsi, B.R.; Chillet, M. 2012. Effects of Black Leaf Streak Disease and Sigatoka Disease on Fruit Quality and Maturation Process of Bananas Produced in the Subtropical Conditions of Southern Brazil. Crop Protection 35:127–131.
- Castelán, F.P.; Abadie, C.; Hubert, O.; Chilin, C.Y.; DeLapeyre, B.L.; Chillet, M. 2013. Relation Between the Severity of Sigatoka Disease and Banana Quality Characterized by Pomological Traits and Fruit Green Life. Crop Protection. 50:61–65.
- Chillet, M.; Hubert, O.; DeLapeyre, L. 2006. Postharvest Disease: Effects of the Physiological Age of Bananas (Musa spp.) on their Susceptibility to Wound Anthracnose due to Colletotrichum musae. Acta Horticulturae. 879: 419–424.
- Chillet, M.; Abadie, C.; Huber, O.; Chilin, C.Y.; DeLapeyre, L. 2009. Sigatoka Disease Reduces the Greenlife of Bananas. Crop Protection. 28: 41–45.
- Chillet, M.; Castelan, F.P.; Abadie, C.; Hubert, O.; Chilin, C.Y.; DeLapeyre, B. L. 2014. Effect of Different Levels of Sigatoka Disease Severity on Banana Pulp Colour and Early Ripening. Canadian Journal of Plant Pathology. 36: 48–53.
- Daniells, J. W.; Lisle, A.T.; Bryde. A. 1994. Effect of Bunch Trimming and Leaf removal at Flowering on Maturity Bronzing, Yield and Others Aspects of Fruit Quality of Bananas in North Queensland. Australian Journal of Experimental Agriculture. 34: 259-265.
- Freeman, B.; Turner, D.W. 1985. The Epicuticular Waxes on the Organs of Different Varieties of Banana (Musa spp.) differ in form, chemistry and concentration. Australian Journal of Botany 33: 393-408.
- Ganry, J.; Meyer, J.P. 1975. Recherche d’une loi d’action de la température sur la croissance des fruits du bananier. Fruits. 30:375–392.
- Guillermet, C.; Le Guen, R.; Dorel, M.; Lescot, T.; DeLapeyre, B. L. 2018. Experimental approaches for agro-ecological management of black leaf streak in dry and humid tropical conditions (en línea). Acta Horticulturae (1196):113–120.
- Guzmán, M.; Marín, D. H.; Pérez, V.; Romero, R.A. 2019. Cultural Control. In: Jones, D.R., editor. Diseases of Banana, Abaca´ and Enset. Wallingford, Oxon OX10 8DE UK: CABI Publishing. 89-95 p.
- Jullien, A.; Chillet, M.; Malezieux, E. 2008. Preharvest Growth and Development Determine Postharvest Green Life of Fruit in Musa (Musa sp. AAA group cv Grande Naine (Cavendish subgroup). Journal Horticultural Science Biotechnology. 83: 506–512.
- Marín, D.H.; Romero, R.A.; Guzmán, M.; Sutton, T.B. 2003. Black Sigatoka: an Increasing Threat to Banana Cultivation. Plant Disease. 87: 208–222.
- Medina, L. 2007. Elaboración de Curvas de Absorción de Nutrimentos en una Plantación Establecida de (Musa sp. AAA), Variedad ¨Williams¨. Tesis M. Sc. Universidad de Costa Rica. San José, Costa Rica. 75 p.
- Mobambo, K.N.; Gauhl, F.; Vuylsteke, D.; Ortiz, R.: Pasberg, G. C.; Swennen, R. 1993. Yield Loss in Plantain from Black Sigatoka Leaf Spot and Field Performance of Resistant Hybrids. Field Crops Research. 35: 35–42.
- Moser, S.; Müller, T.; Holzinger, A.; Lütz, C.; Jockusch, S.; Turro, N.J.; Kräutler, B. 2009. Fluorescent Chlorophyll Catabolites in Bananas Light Up Blue Halos of Cell Death. Proceedings of the National. Academy of Sciences of the United States of America. 106(37):15538–15543. DOI: https://doi.org/10.1073/pnas.0908060106.
- Moser, S.; Müller, T.; Holzinger, A.; Lütz, C.; Kräutler, B. 2012. Structures of Chlorophyll Catabolites in Bananas (Musa acuminata) Reveal a Split Path of Chlorophyll Breakdown in a Ripening Fruit. Chemistry A European Journal. 18(35):10873–10885.
- Ramsey, M.D.; Daniells, J.W.; Anderson, V.J. 1990. Effects of Sigatoka leaf spot (Mycospherella musicola Leach) on Fruit Yield, Field Ripening and Greenlife of Bananas in North Queensland. Science Horticulturae. 41: 305-313
- Robinsom, J.C.; Anderson, T.; Eckstein, K. 1992. The Influence of Functional Leaf Removal at Flower Emergence on Components of Yield and Phothosynthetic Compensation in Banana. Journal of Horticultural Science. 67: 403-420.
- Rodríguez, G.C.; Cayón, S.D.G.; Mira, C.J.J. 2006. Número Mínimo de Hojas Funcionales a Floración y Cosecha en Banano (Musa AAA Simmonds) en Urabá, Colombia. In: Soprano E, Adami TF, Lichtemberg LA, Silva MC, editors. XVII ACORBAT, Brazil, 15–20 octubre 2006, Joinville (Santa Catalina). 319 p.
- Rodríguez, P.; Cayón, G. 2008. Efecto de Mycosphaerella fijiensis sobre la Fisiología de la Hoja de Banano. Agronomía Colombiana. 26:256-265.
- Saborío, G.; Granados, F. 2005. Defoliación Controlada de Plantas Florecidas de Banano (Musa AAA) como Parte de una Estrategia de Manejo Integrado de la Sigatoka negra. Resúmenes Primer Congreso Científico-Técnico Bananero Nacional. Dirección de Investigaciones. Corporación Bananera nacional. Pococí, Costa Rica. 58 p.
- Segura, A.; Rodríguez, W.; Guzmán, G.; González, M. 2005 a. Perfil Fotosintético Foliar y Respuesta a la luz en los Cultivares ¨Grande Naine¨, Valery¨ y ¨Williams¨In: Resúmenes del I Congreso Científico-Técnico Bananero Nacional. Corporación Bananera nacional (CORBANA). Pococí, Limón. Costa Rica. 50 p.
- Segura, A.; Rodríguez, W.; Valle, H.; González, M. 2005 b. Análisis de Crecimiento y Particionamiento de la Biomasa en los Cultivares ¨Grande Naine¨, Valery¨ y ¨Williams¨ In: Resúmenes del I Congreso Científico-Técnico Bananero Nacional. Corporación Bananera nacional (CORBANA). Pococí, Limón. Costa Rica. p 48.
- -egura, A.; Valle, H. 2005. Niveles de Defoliación Controlada a la Floración y su Efecto sobre el Racimo y el Hijo de Sucesión de los Cultivares ¨Grande Naine¨, Valery¨ y ¨Williams. In: Resúmenes del I Congreso Científico-Técnico Bananero Nacional. Corporación Bananera nacional (CORBANA). Pococí, Limón. Costa Rica. 56 p.
- Vargas, A.; Araya, M.; Guzmán, M.; Murillo, G. 2005. Defoliación después de la Floración en Banano del subgrupo Cavendish (Musa AAA). Efecto sobre las Características del Racimo y de la Planta sobre la Severidad de la Sigatoka negra. In: Resúmenes Primer Congreso Científico-Técnico Bananero Nacional. Dirección de Investigaciones. Corporación Bananera nacional. Pococí, Costa Rica. 52 p.
- Vargas, A.; Araya, M.; Guzmán, M. 2006. Efecto de la Defoliación de Plantas de Banano (Musa AAA) a la Floración sobre la Sigatoka Negra /Mycosphaerella fijiensis) y la Producción. Congreso Internacional: Manejo de la Sigatoka Negra en Banano en América latina y el Caribe. Corbana-Inibap-Musalac. San José. Costa Rica. 64 p.
- Vargas, A.; Araya M.; Guzmán M.; Murillo, G. 2009. Effect of Leaf Pruning at Flower Emergence of Banana Plants (Musa AAA) on Fruit Yield and Black Sigatoka (Mycosphaerella fijiensis) disease. International Journal of Pest Management. 55: 19–25.




{kind=link}
{kind=link}
{kind=link}
interesante, muy ilustrativo
Gracias por su comentario, saludos.