
Dr. Carlos De León
Fitopatología, CP
Maíz
a. Resistencia al tizón foliar por turcicum:
(Telomorfo: Trichometasphaeria turcica)
(Anamorfo: Exserohilum turcicum; Sin. Helminthosporium turcicum)
El atizonamiento foliar por turcicum en maíz, una planta alógama, también representa un buen ejemplo de la presencia de razas y la vulnerabilidad de los sistemas de resistencia monogénicos. Mientras que la resistencia poligénica ha sido estable a todas las razas del patógeno a través de los años, en 1960s se determinó la presencia del gene de resistencia Ht, sencillo dominante, que limita la infección en forma de una lesión clorótica. Esta resistencia monogénica se rompió con las nuevas razas 2 y 3 del hongo. De inmediato se encontraron los genes Ht2, Ht3 y HtN con resistencia a éstas razas. El monogene dominante HT2 se detectó en Australia y el HtN se reportó en Suráfrica, presente en la raza Pepitilla. En EEUU, el monogene Ht3 se encontró en un segmento del cromosoma 7 resultado del cruce de Tripsacum floridanum x maíz.

|
Figura 1. Tizón por turcicum en maíz |
b. Resistencia al tizón foliar por maydis:
(Telomorfo: Cochliobolus heterostrophus)
(Anamorfo: Bipolaris maydis. Sin. Helminthosporium maydis
En este hongo se han reportado solamente las razas 0 y T. La resistencia a la raza 0 puede ser monogénica (gene recesivo rhm rhm) o poligénica. La raza T causó gran daño en los años 60s al manifestarse la susceptibilidad de genotipos con androesterilidad citoplásmica T, de Texas (Tcms). En México no representó ningún problema ya que no existen maíces con androesterilidad de ningún tipo.

|
Figura 2. a) Tizón foliar por maydis (Raza 0) y b) Tizón foliar por maydis (Raza T). |
En EEUU y Europa causó cuantiosas pérdidas ya que existe el dominio de la tecnología de híbridos en los que se incorpora la androesterilidad en las líneas que servirán como hembras en la formación de la F1 de los híbridos.
c. Resistencia a roya por polysora (Puccinia polysora):
En los años 40s, se reportó la incidencia de ésta enfermedad en África Oriental en donde era desconocido, presagiando una gran hambruna en esos países. Para los años 50s, Storey había identificado 12 razas fisiológicas del hongo y los correspondientes genes sencillos dominantes que controlaban esas razas, incluyendo los genes Rpp1 al Rpp11. La mayoría de éstos genes se obtuvieron de colecciones de razas de maíz de San Luís Potosí y Michoacán. Al cabo de unos cuantos años, la enfermedad se reportó como controlada y, aun cuando en la actualidad está presente en varias zonas tropicales de esos países, la enfermedad prácticamente no representa ningún problema, por lo que se considera la alternativa de que la enfermedad se controló sin intervención del hombre.
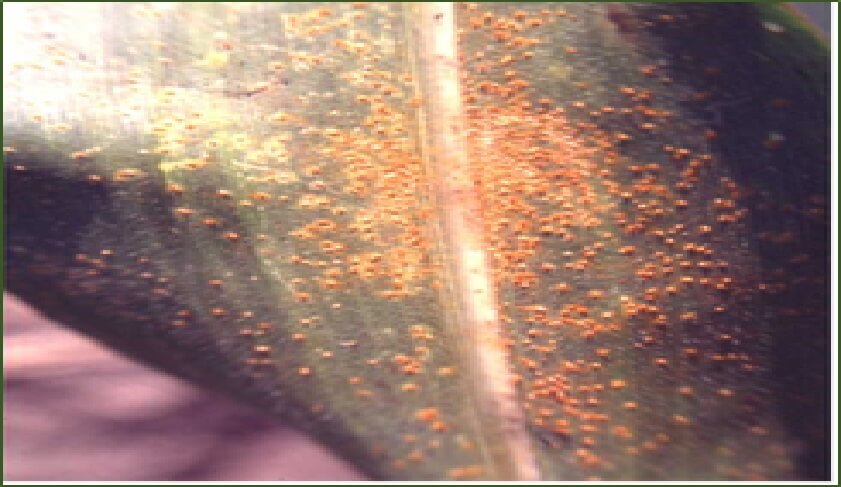
|
Figura 3. Roya por polysora en maíz. |
d. Resistencia a la roya común (Puccinia sorghi):
En los años 50s se reportaron epidemias severas de la enfermedad en varios países. Así, se encontró la resistencia monogénica o específica a razas debida al conjunto de 14 alelos (Rp1-a, Rp1-k, Rp1-b, Rp1-c, etc. en el locus Rp1 de la raza Cuzco Gigante, una raza de maíz harinoso de Perú). El monogene de resistencia Rp1Td se obtuvo de la cruza T. dactyloides x maíz. La forma más común de resistencia es la poligénica o no específica a razas.

|
Figura 4. Roya común del maíz. |
El germoplasma tropical es muy susceptible a ésta enfermedad cuando este se siembra en los valles altos en donde la enfermedad existe en forma endémica. Caso similar se presentó en caso de materiales tropicales que en los años 60s se sembraron en ambientes apropiados para el carbón de la espiga (Sphacelotheca reiliana) como sucedió con cultivares derivados de la raza Tuxpeño que se desarrollaron y sembraron en el Bajío.
e. Resistencia a la pudrición de tallo por Diplodia (Stenocarpella zeae, Sin. Diplodia maydis):
La selección de resistencia se inició en Iowa, EEUU, inoculando con picnidios el primer entrenudo alargado en la base de los tallos y midiendo el área decolorada por la infección en el entrenudo inoculado y la invasión del hongo en nudos adyacentes. La resistencia acumulada por selección recurrente de S1s se midió al final de 3 ciclos de selección comparando las ganancias entre los varios ciclos de selección (C0, C1, C2 y C3). La resistencia a pudrición de tallo y pudrición de mazorca por éste patógeno son controlados en forma independiente.

|
Figura 5. Pudrición de tallo por Stenocarpella. |
f. Achaparramiento (Spiroplasma kunkelii):
En 1961 se inició la selección de resistencia genética en el Campo Cotaxtla del INIA utilizando insectos infectivos del cicadélido Dalbulus maidis, vector del patógeno. Se encontró que la mayoría del germoplasma de maíz tropical originario de México era muy susceptible a la enfermedad. Los materiales que se reportaron como los más resistentes fueron colecciones de Cuba y Rep. Dominicana. Estas actividades se continuaron en el CIMMYT en colaboración con Programas Nacionales de Maíz de Nicaragua y El Salvador. Se determinó que la resistencia seleccionada es poligénica y estable.
En 1973, se reportó que las supuestas razas Río Grande y Mesa Central del hasta entonces conocido como “virus del achaparramiento” eran dos enfermedades diferentes causadas por un espiroplasma y un fitoplasma, respectivamente. Ambos patógenos son trasmitidos, con los virus del rayado fino marafivirus y el virus del enanismo clorótico del maíz (maize chlorotic dwarf virus), por el cicadélido Dalbulus maidis causando lo que se conoce como el complejo del achaparramiento.

|
Figura 6. Achaparramiento del maíz (espiroplasma)
|
g. Pudrición de tallo y mazorca por Fusarium:
[Anamorfo: Fusarium verticillioides (sin. F. moniliforme)]
[Telomorfo: Gibberella moniliforme (sin. G. fujikuroi)]
Se ha desarrollado mucho trabajo en la selección y acumulación en resistencia a éste problema por investigadores de varios países, con pocos resultados positivos. En 1978 se iniciaron en México trabajos para selección de resistencia a estos dos problemas.
- Pudrición de tallo por Fusarium
Aun cuando existen muchos autores que consideran que ésta enfermedad no representa ser problema de importancia económica, se sabe que ésta es común en regiones con ambientes de trópico húmedo con períodos de sequía, comunes en éstas regiones tropicales, aunque se puede encontrar con distribución cosmopolita en todas las regiones productoras de maíz. La identificación de plantas enfermas por éste hongo representa un problema debido a la dificultad de diferenciar sus síntomas en tallos con los producidos por Diplodia y Gibberella. La importancia de la enfermedad está en debate debido a que el hongo F. verticillioides puede aislarse de tallos enfermos y de sanos, por lo que se asume que el hongo está asociado al desarrollo del tallo pero no como parásito. Sin embargo, puede causar lesiones severas en tallos inoculados artificialmente y causar pérdidas económicas en regiones tropicales con períodos de sequía. En el campo, las plantas afectadas se pueden identificar porque estas están caídas presentando un micelio blanquizco en los nudos. El parénquima del tallo desaparece quedando los haces vasculares sueltos con un color café.

|
Figura 7. Pudrición de tallo por F. verticillioides. |
En condiciones de laboratorio, el hongo crece con un micelio blanco, amarillento y en diferentes intensidades de violáceo. Produce abundantes cadenas de microconidias. Es poco común la producción de macronidias. En condiciones de campo es raro encontrar el estado telomorfo (Gibberella moniliforme, sin. G. fujikuroi). Un hongo causando una pudrición de tallo muy similar en maíz es F. moniliforme var. subglutinans que se puede diferenciar del anterior porque éste produce micronidias en falsas cabezas, nunca en cadenas.
Inoculación
Utilizando un método de inoculación artificial apropiada se aumenta la eficiencia de la selección de resistencia y se evitan escapes. Normalmente, se inocula el primer entrenudo alargado con palillos contaminados con el hongo. En trabajos menos críticos se empujan los tallos dándoles un golpe en la base. La evidencia indica que existe una correlación positiva entre la reacción a la inoculación artificial y la incidencia natural, que está también asociada al acame del tallo. Existen resultados positivos publicados utilizando el método de selección recurrente de medios hermanos modificado.
Reacción de resistencia
Se considera que la resistencia a un hongo patógeno del tallo está asociada con resistencia a otros hongos. Sin embargo, la resistencia a un patógeno en estado de plántula, en el tallo o en la mazorca está controlada por caracteres independientes.
Está comprobado que el germoplasma resistente continúa acumulando materia seca por varias semanas después de polinización mientras que los susceptibles detienen su crecimiento en la polinización, seguida rápidamente por el proceso de senescencia. El germoplasma precoz, en general, es más susceptible a la pudrición de tallo que el tardío. También, se ha observado que materiales con alto rendimiento son más susceptibles que aquellos con baja producción.
Caracteres de resistencia
Todos los reportes indican que la resistencia está controlada por caracteres cuantitativos con acción aditiva. Cuando se cruzan algunas líneas, éstas pueden ser más eficientes en transmitir su resistencia que otras con similares características de resistencia, indicando la presencia de efectos no aditivos. En todos los casos, las F1s tienden a ser similares al progenitor más resistente. Reportes de Egipto, indican que la resistencia a F. moniliforme es controlada por 2 genes dominantes, localizados en los brazos cortos del cromosoma 7 y largo del 10.
Trabajos de resistencia con Diplodia maydis reportan que ésta involucra acción génica aditiva, dominancia y a veces epistasis ya que las F1s son más resistentes que cualquiera de los padres. Reportes de India relacionados con resistencia a C. acremonium mencionan que ésta es incompletamente dominante sobre susceptibilidad y los efectos aditivo x aditivo y dominante x dominante no son importantes.
- Pudrición de mazorca por Fusarium
A nivel internacional tal vez esta sea la principal enfermedad del maíz. En condiciones de trópico seco o húmedo, F. verticillioides (sin. F. moniliforme) causa pérdidas de consideración.
Las mazorcas afectadas presentan una pudrición aislada de unos cuantos granos o áreas grandes de granos afectados. El micelio que desarrolla en los granos es color blanco o rosa pálido. Los granos afectados presentan un rayado en la superficie debido a la presencia de micelio bajo el pericarpio.
Inoculación
La inoculación eficiente de mazorcas con F. moniliforme se logra inyectando conidias del hongo en la punta de la mazorca cuando los estigmas empiezan a marchitarse 3-4 días después de la polinización. Para esto se utiliza una jeringa automática. Otros métodos menos eficientes incluyen el introducir un punzón contaminado con conidias del hongo por un lado del jilote recién polinizado o asperjando conidias en los estigmas recién polinizados cubriendo el jilote inoculado con una bolsita de plástico para mantener humedad.

|
Figura 8. Pudrición de mazorca. |
Caracteres de resistencia
La resistencia a pudriciones de mazorca depende de factores como la fecha de siembra, resistencia fisiológica, cubierta de la mazorca y estado de madurez de la planta. El maíz con alta calidad de proteína, ligada al gene homocigótico recesivo opaco 2 (o2 o2), es más susceptible a F. moniliforme que sus contrapartes normales, pero esta reacción puede mejorarse seleccionando genes modificadores a un endosperma más duro.
Resistencia
La mayoría de los trabajos publicados indican que la resistencia a F. moniliforme está controlada por acción génica aditiva con algo de dominancia.
h. Complejo de la mancha de asfalto (Phyllachora maydis, Monographella maydis y Conyothirium phyllachorae):
Los trabajos de selección y acumulación de resistencia han sido desarrollados en México usando infección natural en áreas de alta incidencia de la enfermedad.
Para la acumulación de genes de resistencia se siguió un programa de selección recurrente de S1-S2 que resultaron en dos poblaciones base de maíz, una Amarilla-TSR (Tar Spot Resistant) y otra Blanca-TSR. Se reportó que la resistencia puede ser monogénica o poligénica con pocos genes modificadores involucrados.
i. Míldeus (Peronosclerospora spp.):
Se han reportado 9 especies de mildews afectando maíz, incluyendo una especie en el género Sclerospora, 2 en Sclerophthora y 5 en Peronosclerospora. Las especies más virulentas son P. philippinensis, P. sacchari y P. maydis, presentes en Filipinas, Taiwan e Indonesia, respectivamente.

|
Figura 9. Mancha de asfalto en maíz.. |
En los años 60s se introdujo a EEUU la especie P. sorghi, en semilla de sorgo procedente de África. Esta especie no es muy agresiva. Diez años más tarde, la enfermedad se había distribuido a países en todo el Continente Americano, en donde era desconocida. Selección de maíces a las especies agresivas resultó en resistencia a todas las especies en los 3 géneros. No sucede lo mismo en caso la selección de resistencia se haga a alguna de las especies menos virulentas. La resistencia puede estar controlada por el monogene dominante Dmr, reportado en Taiwan, o por 2-3 genes con acción aditiva.

|
Figura 10. Mildiu en hojas y punta loca en maíz. . |
j. Mosaico enanismo del maíz (MDMV):
Existen varias razas del virus siendo la más común la raza A. La raza B no afecta a pasto Johnson y está considerada como una raza del Sugar Cane Mosaic Virus (SCMV). Por similitudes generales, se considera que el MDMV es una raza del SCMV. La resistencia está controlada por varios genes con acción aditiva.

|
Figura 11. Mosaico de caña de azúcar (SCMV) en maíz. |
Alfalfa
Resistencia al marchitamiento bacteriano (Corynebacterium insidiosum):
La selección de resistencia a la marchitez bacteriana en alfalfa, una planta alógama, se hizo en invernadero siguiendo la técnica del "cotiledón herido", eliminando medio cotiledón después de aplicar el inóculo. La selección de resistencia era en base de plántulas individuales. Se encontró que la resistencia era debida a pocos genes mayores y que también podía deberse a varios genes aditivos con efectos menores.

|
Figura 12. Secciones de tallo de alfalfa con marchitez bacteriana. |




{kind=link}
{kind=link}
{kind=link}
Aún no hay comentarios sobre el artículo